使用本网站需要启用 JavaScript, 请启用后刷新页面获得更好的体验
登录
注册
首页
个人基因检测
临床应用研究
科研合作项目
合作与服务
社区
研究所
微解读
应用
姓氏祖源
基因关系
原始数据
纯合片段
使用 WeGene 需要启用 Cookies, 请启用后刷新页面获得更好的体验
社区首页
祖源讨论区
讨论详情
暴走的MEIS2基因
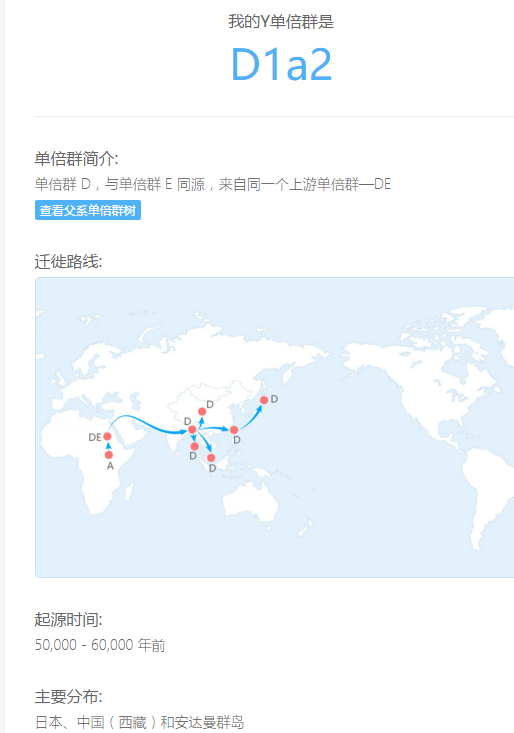
父系单倍群D
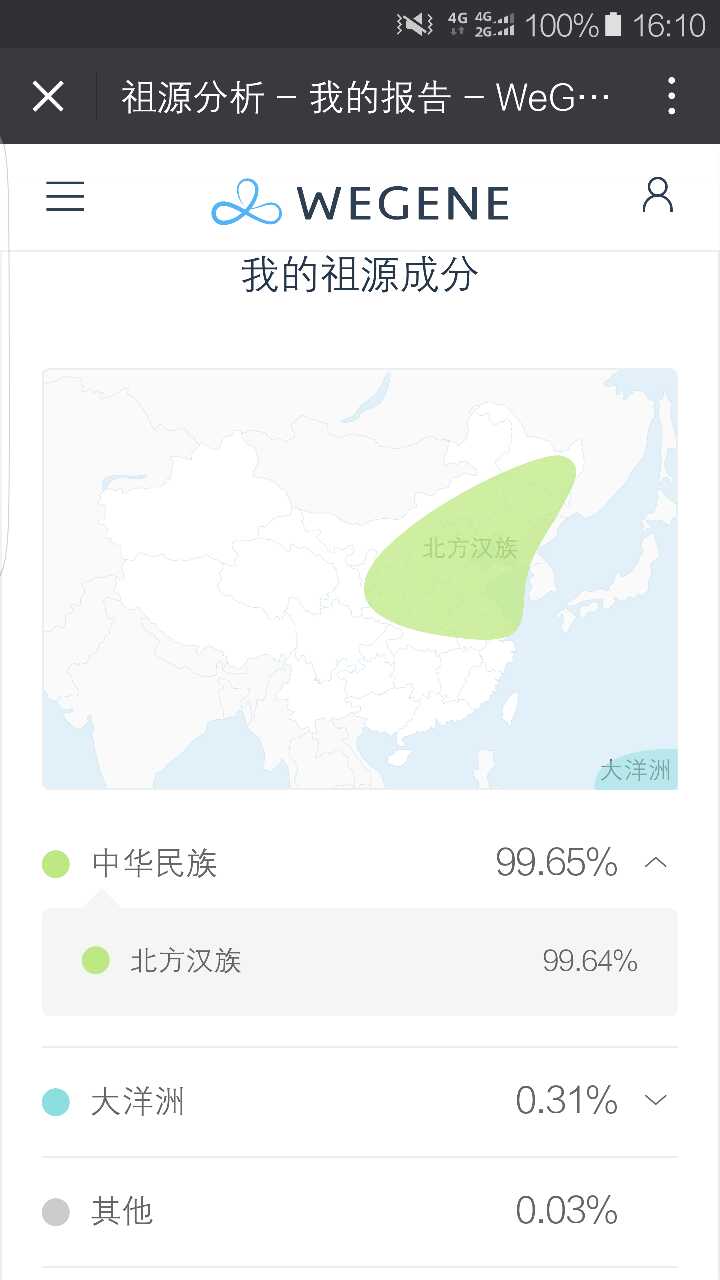
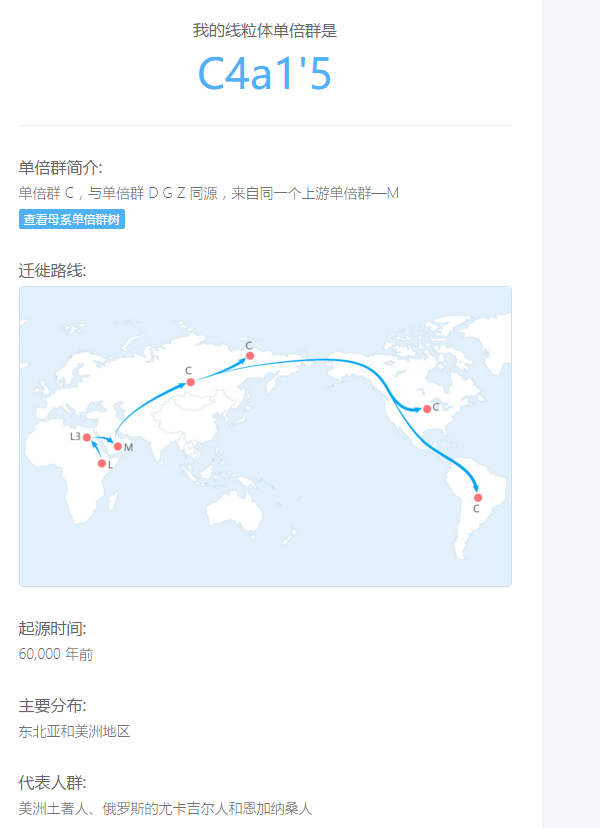
父系D1a2 母系C4a1'5 北方汉族99.64% 求大神分析
求组织 有吗。。。
2017-03-03 • IP属地成都
与内容相关的链接
提交
按热门排序
按默认排序
5 个回复
心软的CDH13基因
严姓有一支出自蜀郡严道(是为阆州严氏),属于以邑为氏。自古秦陇蜀多有交流
匿名用户 • 12 小时前
@清夜
:你说得这个是D3。D1 主要是藏族,羌 彝
-------
楼主是D1a2-P99+,正是老D3,不是老D1。关中早在西汉初期就已多有羌人与汉民混居,后世又挨着西夏故地的党项人聚居区,即便楼主D3不来自蜀地,后世的关中有(老)D3也很正常
暴走的MEIS2基因
这是我这几天找到的一些网上的讨论:
D1高频见于西藏,同时散布东亚,而D2几乎仅局限于日本,D3在西藏。那么,D1是否是旧石器时代留在东亚的残留成分呢?抑或为藏羌族群在历史上与汉族的混血所致?
分析一下17位点的D1数据,藏族(1-35)和苗族(36-39)样本均为纯数字,SNP定型,少量汉族样本为测snp,y-str推测,但基本可以肯定D1。
所有样本(安徽anhui和蒙古Borj两例样本,385双拷贝,未计入)的TMRCA是5290年,95%可信区间3460年-1.78万年。西北马家窑文化的年代为5300-4100年前。吻合度较高。
除了一例山东Shan2样本外,左下角的簇基本上都是藏族,年代为2684年,95%可信区间1753-9039年前。
汉族样本没有出现与藏组样本分离的特征,二是与藏族样本分别聚类,因此,并不能支持汉族的D1属于独特的旧石器残留成分,而当为在近1000-6000年内,与藏族极其先祖相互融合的结果。
又有人不同意如上所说:D1是汉族最初形成时就带有的成分,可能不是来自藏族的。东亚许多民族都有D1,虽然特别低频,D1是最早走出中亚的,融入了不止一个族群,藏缅族群特有的是D3,是最后走出中亚的,几乎只在康巴人安多人和普米族内高频,分布地也未超出藏缅族群。
暴走的MEIS2基因
本人姓严,祖籍陕西宝鸡,求更多大神分析
wang
-
哈佛医学院、德国马普所分子人类学博士后
D1a2-P99:
单倍群D-M174在安达曼尼格利陀人、北部藏缅群体和日本的阿伊努人中高频分布,在其他东亚、东南亚和中亚群体也有低频分单倍群D-M174在安达曼尼格利陀人、北部藏缅群体和日本的阿伊努人中高频分布,在其他东亚、东南亚和中亚群体也有低频分布。D下分D-M15、D-M55和D-P99三个主要支系,还有许多未明确定位的小支系。D1-M15在藏族、羌语支和彝语支人群中广泛分布,在东亚其他群体中也有低频分布。D-M55仅分布于日本,占日本40%以上,是上古绳文人的主要成分。D-P99在青藏高原东部(康区)、白马人及纳西族等群体中高频。D*多在安达曼群岛被发现,且已被隔离了至少2万年。其他一些被包含在D*中的小支系也多分布于西藏周边藏缅语人群、东南亚人群,阿尔泰人中也有少量来源不明的D*。
MrTiung
y染色体似乎很少见,哪里人?
要回复问题请先
登录
或
注册
发起讨论
父系单倍群D
22 个讨论


5 个回复
匿名用户 • 12 小时前
@清夜:你说得这个是D3。D1 主要是藏族,羌 彝
-------
楼主是D1a2-P99+,正是老D3,不是老D1。关中早在西汉初期就已多有羌人与汉民混居,后世又挨着西夏故地的党项人聚居区,即便楼主D3不来自蜀地,后世的关中有(老)D3也很正常
赞同来自: 清夜
D1高频见于西藏,同时散布东亚,而D2几乎仅局限于日本,D3在西藏。那么,D1是否是旧石器时代留在东亚的残留成分呢?抑或为藏羌族群在历史上与汉族的混血所致?
分析一下17位点的D1数据,藏族(1-35)和苗族(36-39)样本均为纯数字,SNP定型,少量汉族样本为测snp,y-str推测,但基本可以肯定D1。
所有样本(安徽anhui和蒙古Borj两例样本,385双拷贝,未计入)的TMRCA是5290年,95%可信区间3460年-1.78万年。西北马家窑文化的年代为5300-4100年前。吻合度较高。
除了一例山东Shan2样本外,左下角的簇基本上都是藏族,年代为2684年,95%可信区间1753-9039年前。
汉族样本没有出现与藏组样本分离的特征,二是与藏族样本分别聚类,因此,并不能支持汉族的D1属于独特的旧石器残留成分,而当为在近1000-6000年内,与藏族极其先祖相互融合的结果。
又有人不同意如上所说:D1是汉族最初形成时就带有的成分,可能不是来自藏族的。东亚许多民族都有D1,虽然特别低频,D1是最早走出中亚的,融入了不止一个族群,藏缅族群特有的是D3,是最后走出中亚的,几乎只在康巴人安多人和普米族内高频,分布地也未超出藏缅族群。
赞同来自:
赞同来自:
单倍群D-M174在安达曼尼格利陀人、北部藏缅群体和日本的阿伊努人中高频分布,在其他东亚、东南亚和中亚群体也有低频分单倍群D-M174在安达曼尼格利陀人、北部藏缅群体和日本的阿伊努人中高频分布,在其他东亚、东南亚和中亚群体也有低频分布。D下分D-M15、D-M55和D-P99三个主要支系,还有许多未明确定位的小支系。D1-M15在藏族、羌语支和彝语支人群中广泛分布,在东亚其他群体中也有低频分布。D-M55仅分布于日本,占日本40%以上,是上古绳文人的主要成分。D-P99在青藏高原东部(康区)、白马人及纳西族等群体中高频。D*多在安达曼群岛被发现,且已被隔离了至少2万年。其他一些被包含在D*中的小支系也多分布于西藏周边藏缅语人群、东南亚人群,阿尔泰人中也有少量来源不明的D*。
赞同来自: 祁墨阳
赞同来自:
要回复问题请先登录或注册